Fertilization Process
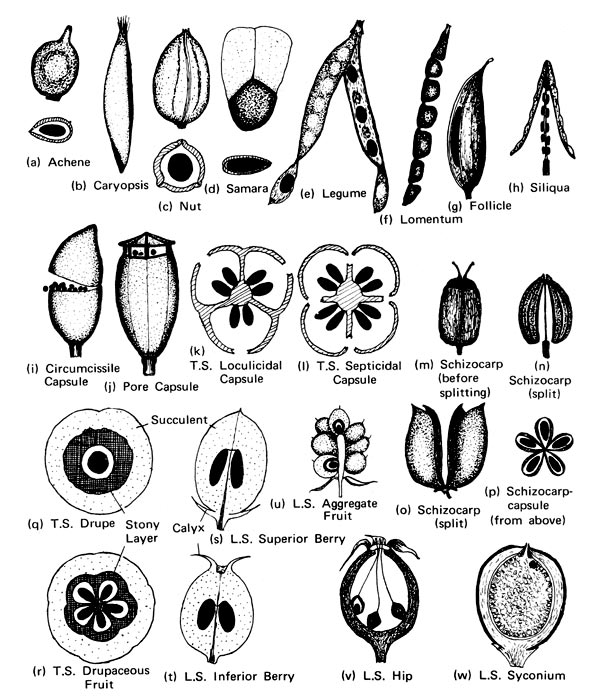
Double fertilization is a complex fertilization mechanism that has evolved in angiosperms (flowering plants) following pollination. It involves the joining of a female gametophyte (embryo sac) with two male gametes (sperm), resulting in the formation of a fruit and its seeds. This process begins when a pollen grain is deposited on the stigma of the flower’s carpel (female reproductive structure). The pollen grain then begins to germinate (requiring water, oxygen and chemical signals to do so), forming a pollen tube that travels down toward the ovary through the style. The tip of the pollen tube then enters the ovary and penetrates the ovule sac through the micropyle. Of the two sperm cells that are released in the megagametophyte, one sperm fertilizes the egg cell, forming a diploid cell (two copies of each chromosome) called a zygote; the other fuses with two polar nuclei (central maternal nuclei), forming a triploid cell that develops into the endosperm, a nutrient-rich tissue which provides nourishment to the developing embryo. The polar nuclei that contribute to the endosperm arises by mitosis from the same single meiotic product that gave rise to the egg. The maternal contribution to the genetic constitution of the triploid endosperm is double that of the embryo. After the completion of this fertilization, no other sperm can enter the ovule. The fertilized ovule then forms the seed, with its outer layers forming the testa, whereas the wall of the ovary becomes the pericarp (fruit wall) which envelop the seeds and may function to disperse them.
Embryonic Development
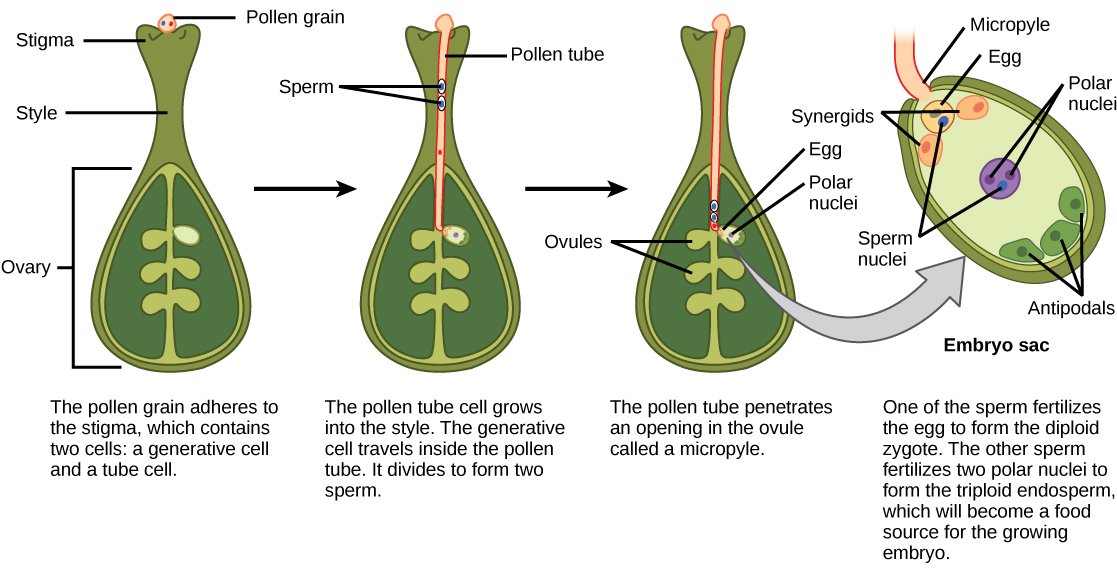
After fertilization, embryonic development begins. The zygote divides to form two cells: the upper cell (terminal cell) and the lower cell (basal cell). The division of the latter results in the apparition of the suspensor that connects the endosperm to the embryo and provides a pathway for nutrition from the mother plant to the growing embryo. The terminal cell also goes under a division which leads to the apparition of the proembryo, a cluster of cells presents in the ovule of the fertilized flowering plant that will then develop into the embryo. In dicots, the developing embryo features a heart shape that indicates the presence of the two rudimentary cotyledons. In non-endospermic dicots, the developing endosperm is eventually digested, then making the two cotyledons the holder of the food reserves. Toward the end of their development, the embryo and cotyledons ultimately fill the seed, which is then ready to be dispersed. Embryonic development is suspended after some time, with growth only resuming when the seed germinates. The developing plant will rely on the cotyledons’ food reserves until the first of leaves start to photosynthesis.
In some plants, seeds or embryos might begin to develop prior to their detachment from the parent; a phenomenon known as vivipary. The newly-formed cormlets or plantlets then fall on the ground in favorable circumstances, conferring them a head start over other fallen seeds. The embryos of such seedlings can develop either through sexual reproduction (such as mangroves) or asexual vegetative reproduction (akin to cloning).
Simple Fruits and Their Types
Simple fruits are the result of the ripening-to-fruit of a simple or compound ovary in a single flower with a single pistil. In contrast, a single flower with numerous pistils typically produces an aggregate fruit upon fertilization; and the merging of several flowers, or a multiple of flowers, results in a multiple fruit. A simple fruit is further classified whether it is dry or fleshy.
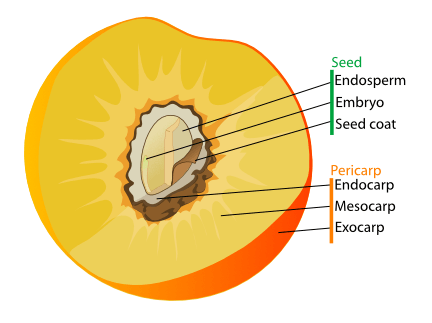
In fleshy fruits, the pericarp is typically made up of three distinct layers: the epicarp (also known as exocarp), which is the outermost layer; the mesocarp, which is the middle layer; and the endocarp, which is the inner layer surrounding the ovary or the seeds. In a citrus fruit, the epicarp and mesocarp make up the peel. In dry fruits, the layers of the pericarp are not clearly distinguishable.
To distribute their seeds, dry fruits may split open, discharge or distribute their seeds through dehiscence and an external abiotic vector (i.e. wind) or indehiscence (decay and degradation of the fruit, or excretion of the fruit by frugivores). Fleshy fruits do not split open and are indehiscent and may also rely on frugivores for distribution of their seeds.
Types of seed dispersal are further discussed in Module I.
Types of dry simple fruits
- achene – most commonly seen in aggregate fruits (strawberry)
- capsule – (Brazil nut)
- caryopsis – (cereal grains, including wheat, rice, oats, barley)
- cypsela – an achene-like fruit derived from the individual florets in a capitulum (dandelion)
- fibrous drupe – (coconut, walnut)
- follicle – follicles are formed from a single carpel, and opens by one suture (milkweed); also commonly seen in aggregate fruits (magnolia, peony).
- legume – (bean, pea, peanut)
- loment – a type of indehiscent legume (sweet vetch or wild potato)
- nut – (beechnut, hazelnut, acorn (of the oak)
- samara – (ash, elm, maple key)
- schizocarp (carrot)
- silique – (radish)
- silicle – (shepherd’s purse)
- utricle – (beet, Rumex)
Types of fleshy simple fruits
- berry – this is the most common type of fleshy fruit, with the entire outer layer of the ovary wall ripening into a potentially edible pericarp.
- stone fruit or drupe – the definitive characteristic of a drupe is the hard, lignified stone (sometimes called pit); derived from the ovary wall of the flower (apricot, cherry, olive, peach, plum, mango).
- pome – pome fruits such as apples, pears, rosehips and saskatoon berry are syncarpous (fused) fleshy fruits, developing from a half-inferior ovary.